


The fruit fly Drosophila has been instrumental in identifying the genes of the Wnt pathway (called Wingless or Wg in the fly). The Naked Cuticle gene (Naked for short or Nkd1/2 in vertebrates) was discovered as a negative feedback regulator of the Wg pathway. This means that Wg induces the expression of Naked, which turns around and attenuates the Wg signal. We have cloned the zebrafish homologues of Naked called Nkd1 and Nkd2. While both are sufficient to inhibit Wnt signaling in vertebrates (Van Raay et al., 2007), our lab focuses on Nkd1 because only Nkd1 is induced by Wnt signaling, at least in zebrafish. Combining the results from work done in Drosophila with our efforts in zebrafish (see left sidebar), we have discovered that Nkd1 needs to be activated by the Wnt ligand in order to be functional. Using the car analogy again, imagine that the Wnt ligand is the driver who controls both the gas pedal and the brakes. This work was the first demonstration that a Wnt ligand was required to activate a negative feedback regulator (Larraguibel et al., 2015).
There is another Wnt negative feedback regulator called Axin2. Axin2 and Nkd1 share many of the same expression domains and together seem to represent universal and obligate targets of the vertebrate Wnt signaling pathway. Axin2 transcription is often used as readout of active Wnt signaling. However, unlike Nkd1, Axin2 does not need to be activated by the Wnt ligand. Our lab will continue to use zebrafish to investigate how Nkd1, Axin2 and other regulators work together to fine tune the Wnt signaling pathway that so elegantly patterns the body plan in the animal kingdom.
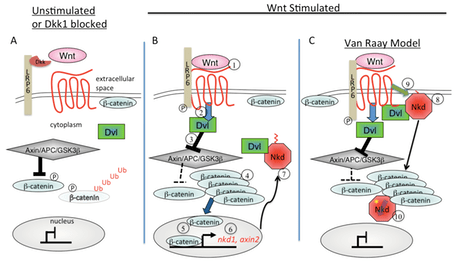
Figure 1. Wnt signaling.
- Simplified model of inactive Wnt signaling. In the absence of a Wnt ligand, or by the presence of a Wnt ligand antagonist such as Dkk1, the destruction complex phosphorylates β-catenin resulting in its ubiquitin mediated degradation.
- In the presence of a Wnt ligand, the destruction complex becomes inactivated resulting in cytoplasmic and nuclear accumulation of β-catenin and the activation of target genes, two of which are nkd1 and axin2.
- Our research has demonstrated that myristoylated Nkd1 localizes to the membrane and is activated by the Wnt ligand-receptor complex. This, in turn, results in a redistribution of Nkd1 away from the membrane into the cytoplasm where it interacts with β-catenin and prevents its nuclear accumulation. This work has all been performed in vivo using zebrafish blastula cells.